![]() ISSN 0798 1015
ISSN 0798 1015

![]() ISSN 0798 1015
ISSN 0798 1015
Vol. 38 (Nº 04) Año 2017. Pág. 3
Sabrina do Couto de MIRANDA 1; Plauto Simão DE-CARVALHO 2; Mercedes Maria da Cunha BUSTAMANTE 3; Manoel Cláudio da SILVA JÚNIOR 4
Recibido: 08/08/16 • Aprobado: 23/08/2016
RESUMO: Este trabalho analisou a variação temporal dos parâmetros estruturais da vegetação lenhosa de cerrado sentido restrito sobre Neossolos Quartzarênicos nos municípios de Correntina e São Desidério, Bahia, Brasil. Foram realizadas amostragens em 1989 e 2010 utilizando parcelas de 1.000 m2 onde os indivíduos lenhosos foram mensurados. No total foram amostradas 95 espécies e 32 famílias. Em 21 anos a biomassa aérea da vegetação lenhosa em Correntina aumentou 3,35 Mg.ha-1 (0,16 Mg.ha-1.ano-1) e em São Desidério o aumento foi de 5,66 Mg.ha-1 (0,27 Mg.ha-1.ano-1). Os cerrados estudados apresentaram pequena variação nos parâmetros florístico-estruturais analisados. |
ABSTRACT: This study aimed to analyze the temporal variation in structural parameters of woody vegetation of cerrado sensu stricto on soils Quartzipsamments in the municipalities of Correntina and São Desidério, Bahia State, Brazil. Two samples were collected, in 1989 and 2010, using plots of 1.000 m2 were all woody individuals were measured. In total we were sampled 95 species and 32 families. In 21 years, the aboveground wood biomass in Correntina increased 3.35 Mg.ha-1 (0.16 Mg.ha-1.year-1) and São Desidério the increase was 5.66 Mg.ha-1 (0.27 Mg.ha-1.year-1). Cerrado on soils Quartzipsamments showed little variation in floristic-structural parameters over time. Key words: woodlands formations, dynamic, woody biomass. |

O cerrado sentido restrito é a fitofisionomias que melhor caracteriza o aspecto savânico do Cerrado e este originalmente ocupava cerca de 70% do bioma (FELFILI; SILVA JÚNIOR, 1993, 2005). Caracteriza-se por árvores e arbustos de pequeno porte, irregularmente ramificados, geralmente tortuosos e com adaptações para sobrevivência à seca e ao fogo (FURLEY, 1999; RIBEIRO; WALTER, 2008). Trata-se de uma comunidade vegetal que apresenta elevada diversidade alfa (FELFILI; SILVA JÚNIOR, 2005). Além disso, nos levantamentos poucas espécies constituem as maiores populações e a distribuição de indivíduos por espécies é desigual ao longo do bioma (FELFILI et al., 2004; FELFILI; SILVA JÚNIOR, 2005).
A maioria dos estudos sobre estrutura da vegetação lenhosa de cerrado sentido restrito foi realizada em comunidades associadas aos Latossolos (NOGUEIRA et al., 2001; ANDRADE et al., 2002; FELFILI et al., 2002, 2004; FONSECA; SILVA JÚNIOR, 2004; FELFILI; SILVA JÚNIOR, 2005; MIRANDA et al., 2007; MIRANDA et al., 2013). Classe de solos mais extensa no Cerrado do Brasil Central que têm por características serem profundos, bem drenados e altamente intemperizados (HARIDASAN, 2005; REATTO; MARTINS, 2005; REATTO et al., 2008).
Portanto, faz-se necessária a condução de estudos que avaliem a estrutura da vegetação de cerrado associado aos outros tipos de solos, tais como os Neossolos Quartzarênicos. Estes solos possuem alto teor de areia e ocupam 14,5% do Cerrado (REATTO et al., 2008). Além disso, são susceptíveis à erosão quando ocorre a retirada da vegetação nativa. (HARIDASAN, 2001). Podem ser encontrados no oeste da Bahia, região que se está sob intensa pressão de mudança de uso da terra (MMA/IBAMA/PNUD, 2009) e onde as paisagens naturais estão sendo rapidamente substituídas por culturas comerciais, principalmente soja e algodão.
A alta pressão antrópica no oeste da Bahia ratifica a realização de estudos que visem compreender a variação estrutural da vegetação de cerrado ao longo do tempo. Neste contexto, este trabalho teve por objetivo analisar a variação temporal dos parâmetros estruturais da vegetação lenhosa de cerrado sentido restrito sobre Neossolos Quartzarênicos.
O projeto Biogeografia do Bioma Cerrado (BIO), coordenado por professores da Universidade de Brasília, amostrou a partir de 1980 áreas de Cerrado em três unidades fisiográficas no Brasil Central (COCHRANE et al., 1985) denominadas Chapada Pratinha, Chapada dos Veadeiros e Chapada do Espigão Mestre do São Francisco (FELFILI et al., 1994). Estas unidades englobam, dentre outras fisionomias, cerrado sentido restrito sobre diferentes tipos de solos, históricos de fogo e pressões antrópicas (MIRANDA, 2012).
Na Chapada do Espigão Mestre do São Francisco o projeto BIO estabeleceu parcelas na Bahia: Correntina, principalmente sobre Neossolos Quartzarênicos; São Desidério e Formosa do Rio Preto sobre Latossolos Vermelho-Amarelo e Neossolos Quartzarênicos. E em Minas Gerais: Parque Nacional Grande Sertão Veredas no município de Formoso sobre Neossolos Quartzarênicos (FELFILI; SILVA JÚNIOR, 2001). Neste trabalho foram reamostradas em 2010 dez parcelas em Correntina e nove em São Desidério, inicialmente estudadas em 1989.
O município de Correntina (13°20’25” S e 44°38’34” W) apresenta altitude média de 586 m, as precipitações médias anuais são na ordem de 1.000 mm e a temperatura média para o período (1986-2009) variou de 21,9° C em junho a 26,6° C em outubro, com média anual de 24,3° C (Dados do INMET). Em São Desidério (12°21’08” S e 44°59’02” W) a altitude média é 756 m, as precipitações médias anuais são na ordem de 1.100 mm e a temperatura média para o período (1986-2009) variou de 22,8° C em junho/julho a 27,7° C em outubro, com média anual de 25,2° C (Dados do INMET).
Os cerrados estudados em Correntina e São Desidério não se localizam em unidades de conservação e estão sob intensa mudança de uso da terra (MMA/IBAMA/PNUD, 2009). As parcelas amostradas localizam-se principalmente em áreas de reserva legal averbadas e observou-se presença de gado, gramíneas exóticas e alta frequência de fogo.
No âmbito do projeto BIO as áreas de cerrado sentido restrito foram amostradas com metodologia padronizada. Em cada localidade foram instaladas parcelas disjuntas de 20 x 50 m (1.000 m2) onde todos os indivíduos lenhosos com diâmetro da base (Db30cm) ≥ 5 cm foram mensurados. O diâmetro foi medido com suta de alumínio e a altura com vara graduada em metros. Considerou-se como altura total a projeção vertical da base do tronco até o ramo mais alto ou folhagem. Neste trabalho foi utilizada a mesma metodologia empregada na primeira amostragem em 1989, facilitando comparação entre os dados.
As famílias botânicas foram classificadas de acordo com o sistema do Angiosperm Phylogeny Group III (APG III, 2009) e os nomes dos autores de todas as espécies conferidos por meio de consultas ao “W3 Tropicos” (http://www.mobot.org). O índice de diversidade de Shannon e Wiener (H’) e uniformidade de Pielou (J’) (MAGURRAN; MCGILL, 2011) foram calculados com o auxílio do programa MVSP (KOVACH, 1993). Os parâmetros fitossociológicos (densidade, frequência e dominância, absolutas e relativas, e Índice de Valor de Importância-IVI) foram calculados de acordo com Müeller-Dombois e Ellenberg (1974). A análise da distribuição de diâmetros e alturas das comunidades, em cada área, foi realizada utilizando-se intervalos entre classes de 5,0 cm e 1,0 m, respectivamente, conforme sugerido por outros estudos facilitando comparações (FELFILI, 2001; FELFILI; REZENDE, 2003; MIRANDA et al., 2013).
Para a comparação entre os anos realizou-se triagem dos dados coletados na primeira amostragem verificando-se sinonímias, mudanças de nomes científicos e famílias botânicas, além disso, foram excluídos das análises os indivíduos identificados até gênero. O turnover temporal de espécies foi calculado através do índice proposto por Brown e Kodric-Brown (1977) discutido em Magurran e McGill (2011). Estes autores propuseram t=b+c/S1+S2, onde b=número de espécies presentes apenas no 1º levantamento; c=número de espécies presentes apenas no 2º levantamento; S1=número total de espécies presentes no primeiro levantamento e S2=número de espécies presentes no segundo levantamento.
A similaridade florística entre as áreas/anos foi avaliada com base nos índices de Sørensen, baseado na presença de espécies (MAGURRAN; MCGILL, 2011), e Czekanowski (KENT; COKER, 1992) com base na densidade, ambos calculados pelo programa MVSP (KOVACH, 1993). Nunes et al. (2002) calcularam em 0,5 a mediana do índice de similaridade de Sørensen nas comparações entre 100 parcelas de cerrado sentido restrito no Distrito Federal, assim foram considerados elevados os valores acima de 0,5. O índice de Czekanowski varia de 0 a 1, e geralmente é apresentado na forma de percentagem (KENT; COKER, 1992), considerando-se elevados os valores acima de 50% (FELFILI; REZENDE, 2003). É importante ressaltar que para as análises de similaridade foram excluídas as espécies raras, ou seja, aquelas representadas por apenas 1 indivíduo.
A biomassa aérea da vegetação lenhosa foi estimada através da equação proposta por Rezende et al. (2006) para o cerrado sentido restrito, B=-0,49129+0,02912*Db2*Ht. Onde: B=biomassa (kg.ind-1), Db=diâmetro da base (cm) e Ht=altura (m).
Para avaliar diferenças significativas (α=0,05) nos parâmetros biométricos da vegetação lenhosa nas áreas entre os anos de amostragem aplicou-se o teste t de Student’s, esta análise foi feita com o programa JMP 9.0.2 (SAS Institute Inc.).
No total foram amostradas 95 espécies distribuídas em 32 famílias (Tabela 1). Nos cerrados sobre Neossolos Quartzarênicos, Fabaceae se destacou como a família mais rica com 18 espécies, seguida por Myrtaceae com oito e Vochysiaceae com sete (Tabela 1).
No primeiro levantamento foram registradas 59 espécies em Correntina e 49 espécies em São Desidério, já no segundo levantamento os valores de riqueza foram 59 e 62 espécies, respectivamente. Em 21 anos, os cerrados estudados em Correntina e São Desidério não apresentaram variação significativa nos valores de riqueza (p > 0,05).
Tabela 1 – Lista florística com os dados de densidade dos indivíduos lenhosos levantados
nas duas amostragens realizadas em Correntina e São Desidério, Bahia, Brasil.
Espécies |
Famílias |
Correntina |
São Desidério |
||
2010 |
1989 |
2010 |
1989 |
||
Acosmium dasycarpum (Vogel) Yakovlev = Leptolobium dasycarpum Vogel |
Fabaceae |
11 |
8 |
11 |
4 |
Aegiphila klotzkiana Cham. |
Lamiaceae |
0 |
0 |
2 |
0 |
Agonandra brasiliensis Miers ex Benth. & Hook. f. |
Opiliaceae |
18 |
5 |
31 |
7 |
Anacardium occidentale L. |
Anacardiaceae |
19 |
25 |
3 |
0 |
Andira cujabensis Benth. |
Fabaceae |
13 |
0 |
2 |
0 |
Andira vermifuga Mart. ex Benth. |
Fabaceae |
18 |
21 |
6 |
2 |
Annona coriacea Mart. |
Annonaceae |
0 |
0 |
0 |
3 |
Annona crassiflora Mart. |
Annonaceae |
1 |
0 |
0 |
0 |
Aspidosperma macrocarpon Mart. |
Apocynaceae |
2 |
1 |
6 |
0 |
Aspidosperma tomentosum Mart. |
Apocynaceae |
25 |
7 |
11 |
41 |
Bowdichia virgilioides Kunth |
Fabaceae |
0 |
5 |
8 |
27 |
Buchenavia capitata (Vahl) Eichler |
Combretaceae |
0 |
0 |
4 |
0 |
Byrsonima affinis W.R. Anderson |
Malpighiaceae |
0 |
0 |
0 |
1 |
Byrsonima coccolobifolia Kunth |
Malpighiaceae |
6 |
7 |
8 |
3 |
Byrsonima pachyphylla A. Juss. |
Malpighiaceae |
53 |
30 |
11 |
30 |
Byrsonima verbascifolia (L.) DC. |
Malpighiaceae |
0 |
5 |
0 |
0 |
Caryocar coriaceum Wittm. |
Caryocaraceae |
2 |
12 |
0 |
6 |
Caryocar cuneatum Wittm. |
Caryocaraceae |
5 |
0 |
23 |
0 |
Casearia sylvestris Sw. |
Salicaceae |
0 |
0 |
0 |
7 |
Chrysophyllum marginatum (Hook. & Arn.) Radlk. |
Sapotaceae |
0 |
0 |
1 |
3 |
Connarus suberosus Planch. |
Connaraceae |
26 |
42 |
22 |
24 |
Couepia grandiflora (Mart. & Zucc.) Benth. ex Hook. f. |
Chrysobalanaceae |
22 |
7 |
2 |
0 |
Dalbergia miscolobium Benth. |
Fabaceae |
0 |
0 |
9 |
20 |
Dimorphandra gardneriana Tul. |
Fabaceae |
5 |
0 |
2 |
0 |
Dimorphandra mollis Benth. |
Fabaceae |
11 |
7 |
3 |
6 |
Diospyros burchellii Hiern |
Ebenaceae |
0 |
0 |
4 |
0 |
Diospyros hispida A. DC. |
Ebenaceae |
0 |
0 |
0 |
10 |
Emmotum nitens (Benth.) Miers |
Icacinaceae |
1 |
1 |
6 |
2 |
Enterolobium gummiferum (Mart.) J.F. Macbr. = Pithecellobium gummiferum Mart. |
Fabaceae |
3 |
2 |
2 |
2 |
Eremanthus glomerulatus Less. |
Asteraceae |
0 |
0 |
0 |
6 |
Eriotheca gracilipes (K. Schum.) A. Robyns |
Malvaceae |
11 |
9 |
6 |
9 |
Eriotheca pubescens (Mart. & Zucc.) Schott & Endl. |
Malvaceae |
0 |
0 |
1 |
0 |
Erythroxylum campestre A. St.-Hil |
Erythroxylaceae |
0 |
1 |
0 |
0 |
Erythroxylum deciduum A. St.-Hil. |
Erythroxylaceae |
4 |
2 |
4 |
1 |
Erythroxylum suberosum A. St.-Hil. |
Erythroxylaceae |
3 |
1 |
1 |
1 |
Eugenia dysenterica DC. |
Myrtaceae |
3 |
1 |
0 |
0 |
Gomidesia lindeniana O. Berg = Myrcia lindeniana (O. Berg) C. Wright |
Myrtaceae |
0 |
40 |
0 |
21 |
Guapira graciliflora (Mart. ex J.A. Schmidt) Lundell |
Nyctaginaceae |
1 |
5 |
12 |
18 |
Guapira noxia (Netto) Lundell |
Nyctaginaceae |
1 |
0 |
0 |
0 |
Hancornia speciosa Gomes |
Apocynaceae |
1 |
2 |
0 |
0 |
Handroanthus ochraceus (Cham.) Mattos |
Bignoniaceae |
25 |
9 |
9 |
0 |
Heisteria ovata Benth. |
Olacaceae |
30 |
2 |
2 |
2 |
Hexachlamys edulis (O. Berg) Kausel & D. Legrand |
Myrtaceae |
0 |
0 |
7 |
0 |
Himatanthus drasticus (Mart.) Plumel |
Apocynaceae |
0 |
0 |
1 |
0 |
Himatanthus obovatus (Müll. Arg.) Woodson |
Apocynaceae |
6 |
4 |
8 |
4 |
Hirtella ciliata Mart. & Zucc. |
Chrysobalanaceae |
1 |
0 |
0 |
0 |
Hirtella glandulosa Spreng. |
Chrysobalanaceae |
0 |
1 |
0 |
0 |
Hirtella gracilipes (Hook. f.) Prance |
Chrysobalanaceae |
36 |
56 |
32 |
37 |
Hymenaea stigonocarpa Mart. ex Hayne |
Fabaceae |
3 |
5 |
2 |
1 |
Kielmeyera coriacea Mart. & Zucc. |
Calophyllaceae |
19 |
23 |
12 |
12 |
Kielmeyera lathrophyton Saddi |
Calophyllaceae |
12 |
9 |
13 |
11 |
Lafoensia pacari A. St.-Hil. |
Lythraceae |
3 |
1 |
0 |
0 |
Machaerium opacum Vogel |
Fabaceae |
2 |
0 |
9 |
4 |
Miconia burchellii Triana = Acinodendron burchellii (Triana) Kuntze |
Melastomataceae |
0 |
0 |
1 |
0 |
Miconia ferruginata DC. |
Melastomataceae |
0 |
0 |
0 |
7 |
Miconia irwinii Wurdack |
Melastomataceae |
0 |
0 |
1 |
0 |
Miconia pohliana Cogn. = Acinodendron pohlianum (Cogn.) Kuntze |
Melastomataceae |
0 |
0 |
1 |
0 |
Mouriri pusa Gardner ex Hook. |
Melastomataceae |
3 |
0 |
6 |
6 |
Myrcia ochroides O. Berg |
Myrtaceae |
6 |
0 |
0 |
0 |
Myrcia sellowiana O. Berg |
Myrtaceae |
98 |
0 |
13 |
0 |
Neea theifera Oerst. |
Nyctaginaceae |
5 |
0 |
0 |
6 |
Ouratea hexasperma (A. St.-Hil.) Baill. = Polyouratea hexasperma (A. St.-Hil.) Tiegh. |
Ochnaceae |
29 |
23 |
8 |
48 |
Ouratea parviflora Engl. |
Ochnaceae |
0 |
1 |
0 |
0 |
Plathymenia reticulata Benth. |
Fabaceae |
8 |
3 |
2 |
2 |
Plenckia populnea Reissek |
Celastraceae |
34 |
5 |
8 |
0 |
Pouteria ramiflora (Mart.) Radlk. |
Sapotaceae |
31 |
57 |
20 |
52 |
Pouteria torta (Mart.) Radlk. |
Sapotaceae |
0 |
1 |
4 |
7 |
Psidium laruotteanum Cambess. |
Myrtaceae |
0 |
1 |
0 |
4 |
Psidium myrsinites Mart. ex DC. |
Myrtaceae |
28 |
4 |
1 |
11 |
Psidium pohlianum O. Berg |
Myrtaceae |
3 |
0 |
9 |
0 |
Pterodon emarginatus Vogel |
Fabaceae |
0 |
0 |
3 |
0 |
Pterodon pubescens (Benth.) Benth. |
Fabaceae |
5 |
6 |
4 |
1 |
Qualea grandiflora Mart. |
Vochysiaceae |
19 |
15 |
19 |
53 |
Qualea parviflora Mart. |
Vochysiaceae |
63 |
20 |
82 |
0 |
Rourea induta Planch. |
Connaraceae |
0 |
4 |
0 |
0 |
Salacia crassifolia (Mart. ex Schult.) G. Don |
Celastraceae |
1 |
0 |
2 |
0 |
Salacia elliptica (Mart. ex Schult.) G. Don |
Celastraceae |
0 |
4 |
0 |
0 |
Salvertia convallariodora A. St.-Hil. |
Vochysiaceae |
6 |
5 |
18 |
21 |
Schefflera macrocarpa (Cham. & Schltdl.) Frodin |
Araliaceae |
0 |
0 |
0 |
1 |
Sclerolobium aureum (Tul.) Baill. = Tachigali aurea Tul. |
Fabaceae |
0 |
5 |
2 |
0 |
Sclerolobium paniculatum Vogel = Tachigali vulgaris L.F. Gomes da Silva & H.C. Lima |
Fabaceae |
39 |
24 |
74 |
99 |
Strychnos pseudoquina A. St.-Hil. |
Loganiaceae |
6 |
15 |
0 |
0 |
Stryphnodendron adstringens (Mart.) Coville |
Fabaceae |
2 |
2 |
3 |
0 |
Stryphnodendron rotundifolium Mart. |
Fabaceae |
0 |
0 |
0 |
1 |
Styrax ferrugineus Nees & Mart. |
Styracaceae |
0 |
1 |
0 |
0 |
Tabebuia aurea (Silva Manso) Benth. & Hook. f. ex S. Moore |
Bignoniaceae |
9 |
7 |
0 |
0 |
Terminalia fagifolia Mart. |
Combretaceae |
17 |
16 |
10 |
23 |
Tocoyena formosa (Cham. & Schltdl.) K. Schum. |
Rubiaceae |
2 |
0 |
0 |
0 |
Vatairea macrocarpa (Benth.) Ducke |
Fabaceae |
16 |
20 |
4 |
3 |
Vochysia cinnamomea Pohl |
Vochysiaceae |
0 |
0 |
1 |
0 |
Vochysia elliptica Mart. |
Vochysiaceae |
0 |
1 |
0 |
0 |
Vochysia gardneri Warm. |
Vochysiaceae |
1 |
0 |
0 |
0 |
Vochysia rufa Mart. |
Vochysiaceae |
18 |
27 |
38 |
46 |
Xylopia aromatica (Lam.) Mart. |
Annonaceae |
0 |
2 |
0 |
0 |
Xylopia sericea A. St.-Hil. |
Annonaceae |
0 |
2 |
0 |
0 |
Total |
- |
851 |
627 |
636 |
718 |
Os índices de diversidade (H’) e equabilidade (J’) calculados para Correntina na primeira amostragem foram, respectivamente, 3,50/0,86 e em São Desidério 3,27/0,84, já na segunda amostragem estes valores foram 3,52/0,86 e 3,51/0,85. Os parâmetros biométricos diversidade e equabilidade não apresentaram variação estatisticamente significativa entre os anos (p > 0,05). O turnover de espécies em Correntina foi de 0,25 e em São Desidério 0,32.
Em Correntina a comunidade lenhosa apresentou na primeira amostragem diâmetro médio de 9,37 cm com coeficiente de variação (CV) de 51%, na segunda amostragem o diâmetro médio foi 9,35 cm (CV=52%), assim ao longo do tempo não houve variação neste parâmetro (t=2,10; p = 0,64). Por outro lado, em São Desidério o diâmetro médio da vegetação lenhosa apresentou aumento significativo entre os anos (t=2,12; p = 0,04), de 8,78 cm (CV=42%) na primeira amostragem para 10,04 cm (CV=60%) na segunda. Com relação à altura média, as duas localidades não apresentaram variação neste parâmetro ao longo do tempo, Correntina 3,70 m (CV=41%) e 3,76 m (CV=33%) (t=2,10; p = 0,89), e São Desidério 3,97 m (CV=39%) e 3,99 m (CV=38%) (t=2,12; p = 0,68), valores referentes a primeira e segunda amostragens, respectivamente.
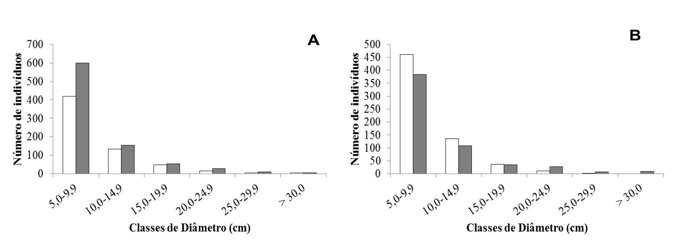
De modo geral, as comunidades apresentaram distribuição dos indivíduos em classes de diâmetro com aspecto J-reverso (Figuras 1A e 1B). A primeira classe diamétrica, com diâmetro da base < 10 cm, abriga cerca de 70% dos indivíduos das comunidades de cerrado. A análise comparativa da distribuição diamétrica entre os anos mostrou que o número de indivíduos na primeira classe (5,0-9,9 cm) aumentou ao longo do tempo no cerrado de Correntina e diminuiu em São Desidério (Figuras 1A e 1B).
Figura 1. Distribuição diamétrica comparativa dos indivíduos lenhosos amostrados em Correntina
(A) e São Desidério (B), Bahia, Brasil. Onde: Barras branca=1ª amostragem (1989); Barras Cinza=2ª amostragem (2010).
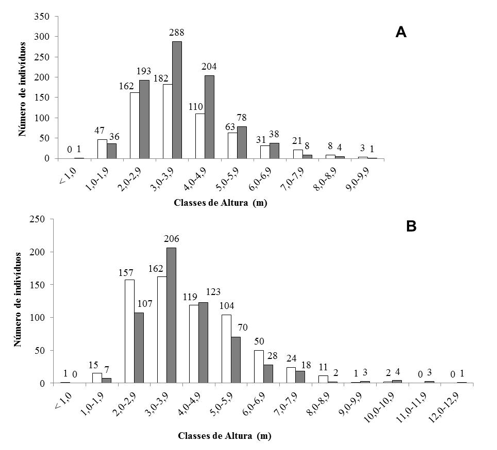
A distribuição das frequências de altura para todos os indivíduos vivos amostrados tendeu à normalidade (Figura 2). Em Correntina, na primeira amostragem, 55% dos indivíduos registrados ocuparam a 3ª e 4ª classes de altura, entre 2,0 e 3,9 metros (Figura 2A). Na segunda amostragem verificou-se que 58% dos indivíduos ocuparam a 4ª e 5ª classes com alturas entre 3,0 e 4,9 metros (Figura 2A). Em São Desidério resultados semelhantes foram observados, na primeira amostragem 50% dos indivíduos ocuparam a 3ª e 4ª classes de altura (Figura 2B) e na segunda amostragem 58% dos indivíduos estavam distribuídos na 4ª e 5ª classes (Figura 2B).
Em Correntina, a densidade de indivíduos vivos foi de 627 ind.ha-1 durante a primeira amostragem e 851 ind.ha-1 na segunda amostragem (Tabela 1). O aumento da densidade lenhosa da comunidade está relacionado ao maior recrutamento de indivíduos nas menores classes de diâmetro (Figura 1A). Por outro lado, em São Desidério a densidade de indivíduos vivos variou de 718 ind.ha-1 na primeira amostragem para 636 ind.ha-1 na segunda amostragem (Tabela 1). A redução na densidade da comunidade se deu nas menores classes diamétricas (Figura 1B).
Os valores de área basal das comunidades lenhosas de Correntina e São Desidério na primeira amostragem foram 5,46 m2.ha-1 e 5,10 m2.ha-1, respectivamente. Já na segunda amostragem os valores foram 7,44 m2.ha-1 em Correntina e 6,84 m2.ha-1 em São Desidério. É importante ressaltar que o aumento na área basal dos cerrados estudados não foi significativo entre os anos (p > 0,05).
Figura 2. Distribuição comparativa das alturas dos indivíduos lenhosos amostrados em Correntina
(A) e São Desidério (B), Bahia, Brasil. Onde: Barras branca=1ª amostragem (1989); Barras Cinza=2ª amostragem (2010).
Do total de espécies amostradas, 31 ocorreram nas duas áreas e nas duas amostragens (Tabela 1). Destas Byrsonima pachyphylla, Connarus suberosus, Hirtella gracilipes, Kielmeyera coriacea, Pouteria ramiflora, Qualea grandiflora, Sclerolobium paniculatum, Terminalia fagifolia e Vochysia rufa foram amostradas com densidade ≥ 10 ind.ha-1 (Tabela 1).
Das espécies amostradas, 35 podem ser consideradas localmente raras, pois ocorreram em apenas uma localidade e em um período de amostragem (Tabela 1). Dentre estas se destacam os representantes do gênero Annona (A. coriacea e A. crassiflora), Diospyros (D. burchellii e D. hispida), Miconia (M. burchellii, M. ferruginata, M. irwinii e M. pohliana) e Xylopia (X. aromatica e X. sericea) (Tabela 1).
Em Correntina, verificou-se que, ao longo de 21 anos, quatro espécies se mantiveram dentre as dez com maiores valores de IVI na comunidade, a saber: B. pachyphylla (6ª posição na 1ª amostragem/4ª posição na 2ª amostragem); H. gracilipes (1ª/3ª posição); Ouratea hexasperma (7ª/10ª posição) e P. ramiflora (2ª/5ª posição).
Em São Desidério, em 21 anos seis espécies se mantiveram dentre as dez espécies com maiores valores de IVI na comunidade, são elas: H. gracilipes (2ª posição na 1ª amostragem/3ª posição na 2ª amostragem); P. ramiflora (4ª/9ª posição); Q. parviflora (3ª/1ª posição); Salvertia convalarieodora (9ª/5ª posição); S. paniculatum (1ª/2ª posição) e V. rufa (6ª/8ª posição).
Durante a primeira amostragem, em Correntina os indivíduos mortos em pé ocorreram em 90% das parcelas com densidade de 40 ind.ha-1. Na segunda amostragem foram encontrados indivíduos mortos em pé em 100% das parcelas de Correntina e a densidade aumentou para 178 ind.ha-1. Em São Desidério, os indivíduos mortos em pé foram registrados em 100% das parcelas durante as duas amostragens (1989 e 2010), os valores de densidade foram 71 ind.ha-1 e 73 ind.ha-1, respectivamente.
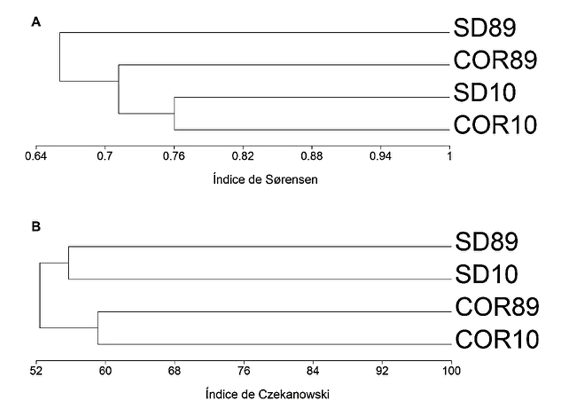
Os índices de similaridade revelaram semelhanças florísticas e estruturais entre as áreas, pois todos os valores de Sørensen ficaram acima de 0,67 (Figura 3A) e de Czekanowski acima de 52% (Figura 3B). A integridade estrutural nas localidades foi mantida ao longo do tempo, porém é possível observar variação na composição de espécies entre os anos (Figura 3A).
Com relação à biomassa aérea da vegetação lenhosa (BAVL), na primeira amostragem os valores estimados para Correntina e São Desidério foram, respectivamente, 9,41 Mg.ha-1 (CV=65,6%) e 8,76 Mg.ha-1 (CV=61,3%). Na segunda amostragem, a BAVL foi estimada em 12,76 Mg.ha-1 (CV=52,5%) em Correntina e 14,42 Mg.ha-1 (CV=87,3%) em São Desidério. A BAVL aumentou 3,35 Mg.ha-1 em Correntina com taxa de incremento de 0,16 Mg.ha-1.ano-1 valor inferior ao encontrado em São Desidério onde o aumento foi de 5,66 Mg.ha-1 com taxa de incremento de 0,27 Mg.ha-1.ano-1. É importante destacar que as diferenças entre os anos em cada área não foram significativas (p > 0,05).
Figura 3. Dendrogramas de classificação da vegetação lenhosa amostrada em Correntina e São Desidério produzidos a partir do índice de Sørensen
(A) e Czekanowski (B). Onde: 89 e 10 são os anos de amostragem (1989 e 2010); COR=Correntina; SD=São Desidério.
A análise dos dados coletados em Correntina e São Desidério mostrou que riqueza e diversidade não variaram significativamente ao longo do tempo. A semelhança florística entre as áreas foi corroborada pelos altos valores do índice de Sørensen. O turnover de espécies nos cerrados sobre Neossolos Quartzarênicos foi baixo, em torno de 30%, contudo este percentual de variação foi detectado na similaridade de Sørensen que mostrou variação entre área/ano de amostragem.
Do ponto de vista estrutural, os cerrados sobre Neossolos Quartzarênicos apresentaram, em média, densidade < 800 ind.ha-1 e área basal < 7 m2.ha-1. Estes parâmetros distinguem estas áreas dos cerrados sobre Latossolos onde os valores médios de densidade e área basal são acima de 1.000 ind.ha-1 e 8 m2.ha-1, respectivamente (MIRANDA, 2012). Ao longo do tempo, os cerrados de Correntina e São Desidério não apresentaram variação significativa em densidade e dominância. Em 21 anos observou-se que a integridade estrutural em cada área foi mantida, fato ratificado pela análise de Czekanowski.
As distribuições de frequência nas classes diamétricas para as áreas de cerrado apresentaram elevada concentração de indivíduos nas menores classes e redução acentuada em direção às maiores. Com base nestas características as comunidades podem ser consideradas auto regenerativas, pois o recrutamento é maior que a mortalidade (FELFILI; SILVA JÚNIOR, 1988; FELFILI, 2001; MIRANDA et al., 2007).
Os cerrados amostrados em Correntina e São Desidério não estão protegidos de distúrbios diversos como queimadas e pisoteio por gado. Além disto, não existem registros da frequência de fogo para as áreas estudadas. Contudo, com base em imagens de satélite MODIS para o período de 2002-2010, Araújo et al. (2012) classificaram a área queimada dos municípios brasileiros em dez categorias que variaram de < 100 km2 (classe 1) a > 15.000 km2 (classe 10). Em um contexto regional, Correntina e São Desidério foram incluídos na 8ª e 9ª classes definidas pelos autores, respectivamente. Assim, podemos assumir que queimadas são frequentes nestas localidades e um forte indício disto é a alta frequência e densidade de indivíduos mortos em pé amostrados. Contudo, com base nas observações em campo tem-se que as parcelas em cada área não são queimadas homogeneamente.
A distribuição das alturas mostrou que os indivíduos lenhosos das duas comunidades apresentaram tendência a maior investimento neste parâmetro biométrico. Acredita-se que este resultado seja uma resposta da vegetação às queimadas frequentes nas áreas. Indivíduos mais altos tendem a escapar do efeito direto das chamas (MIRANDA et al., 2004). A linha de queimada geralmente ocorre entre 1,0 e 1,5 m (COUTINHO, 1990; SAN JOSÉ; FARIÑAS, 1983, 1991), portanto indivíduos com altura acima de 2 metros conseguem proteger suas gemas e ramos jovens aumentado a sobrevivência pós-fogo.
Nos cerrados sobre Neossolos Quartzarênicos poucas espécies foram representativas em abundância e dominância, e a maioria é rara. Nas duas amostragens, as dez espécies com maiores IVI representaram mais de 50% da densidade e área basal das comunidades. Este tipo de distribuição é comumente encontrada em cerrado sentido restrito no Brasil Central (FELFILI et al., 1994, 2000, 2004).
Apenas duas espécies, Hirtella gracilipes e Pouteria ramiflora, foram consideradas representativas dos cerrados sobre Neossolos Quartzarênicos. Estas espécies nas duas amostragens foram comuns às áreas dentre as dez com maiores IVI. De acordo com Ratter et al. (2003), P. ramiflora compõe a lista das 38 espécies amplamente distribuídas no cerrado, já H. gracilipes foi amostrada somente em oito das 315 áreas core estudadas por estes autores, portanto com distribuição mais restrita.
Em 21 anos, os cerrados sobre Neossolos Quartzarênicos apresentaram pequena variação nos parâmetros florístico-estruturais. Outros estudos de longa duração realizados em cerrado sentido restrito corroboram que a vegetação lenhosa tende a manter sua composição florística ao longo do tempo, mesmo sob frequência moderada de distúrbios como o fogo (FELFILI et al., 2000; LIBANO; FELFILI, 2006).
Para as estimativas de biomassa aérea lenhosa tendência similar foi encontrada, não houve variação significativa ao longo do tempo. Este é um importante resultado principalmente porque a maioria das parcelas foi instalada em reservas legais averbadas. De acordo com o Código Florestal (Lei 12.651/2012) trata-se de “área localizada no interior de uma propriedade ou posse rural, com a função de assegurar o uso econômico de modo sustentável dos recursos naturais do imóvel rural, auxiliar a conservação e a reabilitação dos processos ecológicos e promover a conservação da biodiversidade, bem como o abrigo e a proteção de fauna silvestre e da flora nativa”.
Neste sentido, mesmo não estando protegidas de distúrbios como fogo e pisoteio por gado, estas áreas foram fundamentais para a manutenção da biodiversidade e, consequentemente, dos serviços ambientais prestados pela vegetação como manutenção dos estoques de carbono. Assim, deve-se estimular o registro e manutenção das áreas de reserva legal, principalmente no Oeste da Bahia, região sob intensa mudança de uso da terra (MMA/IBAMA/PNUD, 2009).
As áreas de cerrado sentido restrito sobre Neossolos Quartzarênicos, em 21 anos, apresentaram pequena variação nos parâmetros florístico-estruturais avaliados. Estes dados corroboram que a vegetação lenhosa de cerrado tende a manter sua composição florística, mesmo sob frequência moderada de distúrbios (fogo e pisoteio pelo gado).
Os dados da dinâmica temporal de cerrado sobre Neossolos Quartzarênicos são inéditos e revelaram importantes características da vegetação, assim sugere-se a continuação de estudos desta natureza.
ANDRADE, L.A.Z.; FELFILI, J.M.; VIOLATTI, L. “Fitossociologia de uma área de cerrado denso na RECOR-IBGE, Brasília-DF”. In: Acta botânica brasílica, 2002, vol16, n.2, p.225-240.
ANGIOSPERM PHYLOGENY GROUP (APG) III. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. In: Botanical Journal of the Linnean Society, 2009, vol 161, p.105-121.
ARAÚJO, F. M.; FERREIRA, L. G.; ARANTES, A. E. “Distribution patterns of burned areas in the Brazilian biomes: an Analysis based on satellite data for the 2002-2010 period”. In: Remote Sens. 2012, vol 4, p.1929-1946.
COCHRANE, T. T.; SANCHEZ, L. G.; PORRAS, J. A; GARVER, C. L. Land in tropical America. CIAT-EMBRAPA-CPAC, Cali. (3 vols.), 1985.
COUTINHO, L. M. Fire in the ecology of the Brazilian Cerrado. In: HUNTLEY, B. J.; WALKER, B. H. (eds.). Ecology of tropical savannas. Springer-Verlag, Berlim,1990. P. 273-291.
FELFILI, J. M. Distribuição de diâmetros de quatro áreas de cerrado sensu stricto na Chapada do Espigão Mestre do São Francisco. In: FELFILI, J.M.; SILVA JÚNIOR, M.C. Biogeografia do bioma Cerrado: estudo fitofisionômico da Chapada do Espigão mestre do São Francisco. Brasília: Universidade de Brasília, Departamento de Engenharia Florestal. 2001.152p.
FELFILI, J. M.; HARIDASAN, M.; MENDONÇA, R. C.; FILGUEIRAS,T. S.; SILVA JÚNIOR, M. C.; REZENDE, A. V. “Projeto biogeografia do bioma Cerrado: vegetação e solo”. In: Caderno de Geociências, 1994, vol 12, p.75-166.
FELFILI, J. M.; NOGUEIRA, P. E.; SILVA JÚNIOR, M. C.; MARIMON, B. S.; DELITTI, W. B. C. “Composição florística e fitossociologia do cerrado sentido restrito no município de Água Boa-MT”. In: Acta botânica brasílica, 2002, vol16, n.1, p.103-112.
FELFILI, J. M.; REZENDE, R. P. Conceitos e métodos em fitossociologia. Universidade de Brasília, Departamento de Engenharia Florestal. 2003. 68p.
FELFILI, J. M.; REZENDE, A.; SILVA JÚNIOR, M. C.; SILVA, M. “Changes in the floristic composition of cerrado sensu stricto in Brazil over a nine-year period”. In: Journal of Tropical Ecology, 2000, vol 16, p.579-590.
FELFILI, J. M.; SILVA JÚNIOR, M. C. Distribuição dos diâmetros numa faixa de cerrado na Fazenda Água Limpa (FAL) em Brasília-DF. In: Acta botânica brasílica, 1988, vol 2, n.1-2, p. 85-104.
FELFILI, J. M.; SILVA JÚNIOR, M. C. A comparative study of cerrado (sensu stricto) vegetation in Central Brazil. In: Journal of Tropical Ecology, 1993, vol 9, p. 277-289.
FELFILI, J. M.; SILVA JÚNIOR, M. C. Biogeografia do bioma Cerrado: estudo fitofisionômico da Chapada do Espigão Mestre do São Francisco. Universidade de Brasília, Faculdade de Tecnologia, Departamento de Engenharia Florestal. 2001.
FELFILI, J. M.; SILVA JÚNIOR, M. C. Diversidade alfa e beta no cerrado sensu stricto, Distrito Federal, Goiás, Minas Gerais e Bahia. Pp.143-154. In: SCARIOT, A.; SOUSA-SILVA, J. C.; FELFILI, J. M. Cerrado: Ecologia, Biodiversidade e Conservação. Brasília: Ministério do Meio Ambiente. 2005.
FELFILI, J. M.; SILVA JÚNIOR, M. C.; SEVILHA, A. C.; FAGG, C. W.; WALTER, B. M. T.; NOGUEIRA, P. E.; REZENDE, A. V. “Diversity, floristic and structural patterns of cerrado vegetation in Central Brazil”. In: Plant Ecology, 2004, vol 175, p.37-46.
FONSECA, M. S.; SILVA JÚNIOR, M. C. “Fitossociologia e similaridade florística entre trechos de cerrado sentido restrito em interflúvio e em vale no Jardim Botânico de Brasília, DF”. In: Acta botânica brasílica, 2004, vol 18, n.1, p.19-29.
FURLEY, P. A. “The nature and diversity of neotropical savanna vegetation with particular reference to the Brazilian cerrados”. In: Global Ecology and Biogeography, 1999, vol 8, p.223-241.
HARIDASAN, M. 2001. Solos. In: FELFILI, J. M.; SILVA JÚNIOR, M.C. (org). Biogeografia do bioma Cerrado: Estudo fitofisionômico na Chapada do Espigão Mestre do São Francisco. Brasília: Universidade de Brasília, Faculdade de Tecnologia, Departamento de Engenharia Florestal.
HARIDASAN, M. Competição por nutrientes em espécies arbóreas do cerrado. In: SCARIOT, A.; SOUSA-SILVA, J. C.; FELFILI, J. M. (Orgs.). Cerrado: ecologia, biodiversidade e conservação. Brasília: Ministério do Meio Ambiente, 2005. p. 169-178.
KENT, M.; COKER, P. Vegetation description and analysis: a practical approach. Belhaven Press: London. 1992. 363p.
KOVACH, W.L. MVSP – Multivariate Statistical Package, version 3.1. Kovach Computing Services, Pentraeth. 1993.
LIBANO, A. M.; FELFILI, J. M. “Mudanças temporais na composição florística e na diversidade de um cerrado sensu stricto do Brasil Central em um período de 18 anos (1985-2003)”. In: Acta botanica brasilica, 2006, vol 20, p. 927-936.
MAGURRAN, A. E.; MCGILL, B. J. Biological Diversity: frontiers in measurement and assessment. Oxford University Press. 345p. 2011.
MIRANDA, S. C. Variação espacial e temporal da biomassa vegetal em áreas de Cerrado. Tese de doutorado, departamento de Ecologia, Universidade de Brasília. 2012. 142p.
MIRANDA, S. C.; SILVA JÚNIOR, M. C.; SALLES, L. A. “A comunidade lenhosa de cerrado rupestre na Serra Dourada, Goiás”. In: Heringeriana, 2007, vol 1, p. 43-53.
MIRANDA, H. S.; SATO, M. N.; ANDRADE, S. M. A.; HARIDASAN, M.; MORAIS, H. C., Queimadas de Cerrado: caracterização e impactos. In: AGUIAR, L. M. S.; CAMARGO, A. J. A. (eds.). Cerrado: ecologia e caracterização. Embrapa Cerrado, Brasília-DF. p. 69-123. 2004.
MIRANDA, S. C.; SILVA JÚNIOR, M. C.; DE-CARVALHO, P. S. O efeito da proteção do fogo na estrutura da vegetação lenhosa de uma área de cerrado sentido restrito no Brasil Central. In: Heringeriana, 2013, vol 7, n.1, p. 61-72.
MMA, IBAMA, PNUD. Relatório técnico de monitoramento do desmatamento no bioma cerrado, 2002 a 2008: Dados Revisados. Acordo de Cooperação Técnica. Brasília-DF. 2009.
NOGUEIRA, P. E.; FELFILI, J. M.; SILVA JÚNIOR, M. C.; DELITTI, W.; SEVILHA, A. “Composição florística e fitossociologia de um cerrado sentido restrito no município de Canarana-MT”. In: Boletim Herbário Ezechias Paulo Heringer, 2001, vol 8, p. 28-43.
MUELLER-DOMBOIS, D.; ELLENBERG, H. Aims and methods of vegetation ecology. New York: Wiley. 1974. 547p.
NUNES, R. V.; SILVA JÚNIOR, M. C.; FELFILI, J. M.; WALTER, B. M. T. “Intervalos de classe para a abundância, dominância e frequência do componente lenhoso do cerrado sentido restrito no Distrito Federal”. In: Revista Árvore, 2002, vol 26, n.2, p.173-182.
RATTER, J. A.; BRIDGEWATER, S.; RIBEIRO, J. F. “Analysis of the floristic composition of the Brazilian cerrado vegetation III: comparison of the woody vegetation of 376 areas”. In: Edinburgh Journal of Botany, 2003, vol 60, p. 57-109.
REATTO, A.; MARTINS, E. S. Classes de solo em relação aos controles da paisagem do bioma Cerrado. Pp. 49-59. In: SCARIOT, A.; SOUSA-SILVA, J. C. & FELFILI, J. M. (orgs.). Cerrado: Ecologia, Biodiversidade e Conservação. Brasília: Ministério do Meio Ambiente. 2005.
REATTO, A.; COREIA, J. R.; SPERA, S. T. & MARTINS, E. S. Solos do bioma Cerrado: aspectos pedológicos. In: SANO. S. M.; ALMEIDA. S. P.; RIBEIRO. J. F. (eds.). Cerrado: ecologia e flora. Embrapa Cerrados. Brasília-DF. 2008.
REZENDE, A.; VALE, A.; SANQUETTA, C.; FILHO, A. F.; FELFILI, J.M. “Comparação de modelos matemáticos para estimativa do volume, biomassa e estoque de carbono da vegetação lenhosa de um cerrado sensu stricto em Brasília, DF”. In: Scientia Forestalis, 2006, vol 71, p. 65-76.
RIBEIRO. J. F.; WALTER, B. M. T. As principais fitofisionomias do bioma Cerrado. In: SANO, S. M.; ALMEIDA, S. P.; RIBEIRO, J. F. (eds.). Cerrado: ecologia e flora. Embrapa Cerrados. Brasília-DF, 2008, p. 153-212.
SAN JOSÉ, J. J.; FARIÑAS, M. R. “Temporal changes in the structure of a Trachypogon savanna, Venezuela”. In: Ecology, 1983, vol 64, p. 447-453.
SAN JOSÉ, J. J.; FARIÑAS, M. R. “Changes in tree density and species composition in a protected Trachypogon savanna protected for 25 years”. In: Acta Oecologia, 1991, vol 12, p. 237-247.
1. Universidade Estadual de Goiás, Campus Palmeiras de Goiás. Bolsista do Programa de Bolsa de Incentivo à Pesquisa e Produção Científica da UEG (PROBIP); Autora para correspondência. Email: sabrinac.miranda@gmail.com
2. Universidade Estadual de Goiás, Campus Palmeiras de Goiás
3. Universidade de Brasília, Departamento de Ecologia
4. Universidade de Brasília, Centro de Referência em Conservação da Natureza e Recuperação de Áreas Degradadas (CRAD).