![]() ISSN 0798 1015
ISSN 0798 1015

![]() ISSN 0798 1015
ISSN 0798 1015
Vol. 38 (Nº 42) Año 2017. Pág. 5
Laís Araújo FAUSTINO 1; Elisa Maria Gomes da SILVA 2; Lino Roberto FERREIRA 3; Gustavo Antônio Mendes PEREIRA 4; Evander Alves FERREIRA 5; Guilherme Pereira QUEIROZ 6; Douglas Teixeira SARAIVA 7
Recibido: 11/04/2017 • Aprobado: 05/05/2017
RESUMO: Eucalipto em competição com plantas daninhas têm suas características fisiológicas normalmente alteradas. O objetivo desta pesquisa é avaliar o efeito do período de convivência de plantas de Commelina benghalensis nas características fisiológicas do eucalipto. A convivência variou em tempos de 0 a 105 dias. A C. benghalensis afeta o consumo de CO2, o carbono interno, a condutância estomática, a pressão de vapor subestomática, a taxa de transpiração e fotossintética e a eficiência do uso da água do eucalipto. |
ABSTRACT: Eucalyptus in competition with weeds have their physiological characteristics normally altered. The objective of this research is to evaluate the effect of the coexistence period of Commelina benghalensis plants on the physiological characteristics of eucalyptus. The coexistence varied in times of 0 to 105 days. C. benghalensis affects CO2 consumption, internal carbon, stomatal conductance, sub-static vapor pressure, transpiration and photosynthetic rate, and eucalyptus water use efficiency. |

A área ocupada por plantações florestais, no ano de 2014, totalizou 7,74 milhões de hectares, dos quais 71,9% correspondem às plantações de Eucalyptus e 20,5% as de Pinus (IBA, 2015), ainda com grande potencial de crescimento.
É de fundamental importância que os plantios de eucalipto apresentem taxas adequadas de produtividade de madeira, o que é possível com o controle dos fatores que limitam o crescimento e desenvolvimento das plantas de eucalipto (Silva, 1993). Dentre os fatores limitantes, destaca-se a presença e a consequente interferência das plantas daninhas que tem sido considerada um dos maiores problemas na implantação, manutenção e reforma das plantações de eucalipto (Cruz et al., 2010).
As relações competitivas entre plantas são bastante complexas, ocorrendo interação entre as espécies abaixo e acima do solo (Zanine e Santos, 2004). O potencial competitivo das plantas pelos recursos do meio varia em função da espécie presente na área (Rigoli et al., 2008; Silva et al., 2013), do nível populacional (Vidal et al., 2004), da época da emergência em relação à cultura (Silva e Silva, 2013) e das características competitivas dos cultivares (Galon et al., 2007; Fleck et al., 2006).
Quando plantas estão submetidas à alta competição, as suas características fisiológicas normalmente são alteradas, o que resulta em diferenças no aproveitamento dos recursos do ambiente, principalmente no uso da água, que influencia de modo direto a disponibilidade de CO2 no mesofilo foliar e a temperatura da folha e, consequentemente, a eficiência fotossintética da planta (Concenço et al., 2007).
A espécie Commelina benghalensis, conhecida vulgarmente como trapoeraba, tem sido descrita como problemática em plantações de eucalipto pela ampla distribuição e dificuldade de controle, causando prejuízos econômicos às diversas culturas (Rodrigues et al., 2010). A dificuldade de controle se deve, provavelmente, à seleção ocorrida em função da utilização contínua e inalterada dos mesmos métodos de controle e herbicidas (Costa et al., 2004). Em áreas onde o glyphosate tem sido utilizado com frequência, observa-se o aumento da população de C. benghalensis, devido à tolerância a esse herbicida (Rocha et al., 2007; Webster e Grey, 2008).
A maioria dos estudos em que se determinaram os efeitos da competição entre as plantas daninhas e as culturas tiveram como objetivo avaliar apenas o efeito da competição de plantas daninhas na produtividade e/ou no crescimento (acúmulo de biomassa) das culturas, portanto, há uma escassez de estudos envolvendo as aptidões fisiológicas específicas de cada planta.
Diante do exposto, objetivou-se avaliar efeito do período de convivência de plantas de C. benghalensis nas características fisiológicas em duas fases do crescimento das plantas de eucalipto.
O experimento foi conduzido no período de 18 de fevereiro a 03 de junho de 2014, na Estação Experimental Diogo Alves de Melo, Campus da Universidade Federal de Viçosa.
O delineamento experimental utilizado foi em blocos casualizados (DBC) com quatro repetições, no esquema de parcelas subdivididas no tempo onde as parcelas eram constituídas de épocas de avaliação do eucalipto (primeira avaliação/fase de crescimento 1 (30 dias após o transplantio) - segunda avaliação/fase de crescimento 2 (150 dias após o trasplantio)) e as subparcelas de períodos de convivência da cultura com a trapoeraba (0; 0-30; 30-45; 45-60; 60-75; 75-105 dias).
As unidades experimentais foram constituídas por vasos de 110 dm3 de solo em campo, com uma planta de eucalipto e três de C. benghalensis, e irrigadas de modo que mantivessem adequada disponibilidade de água, acompanhando as variações climáticas durante a época de condução (Figura 1).
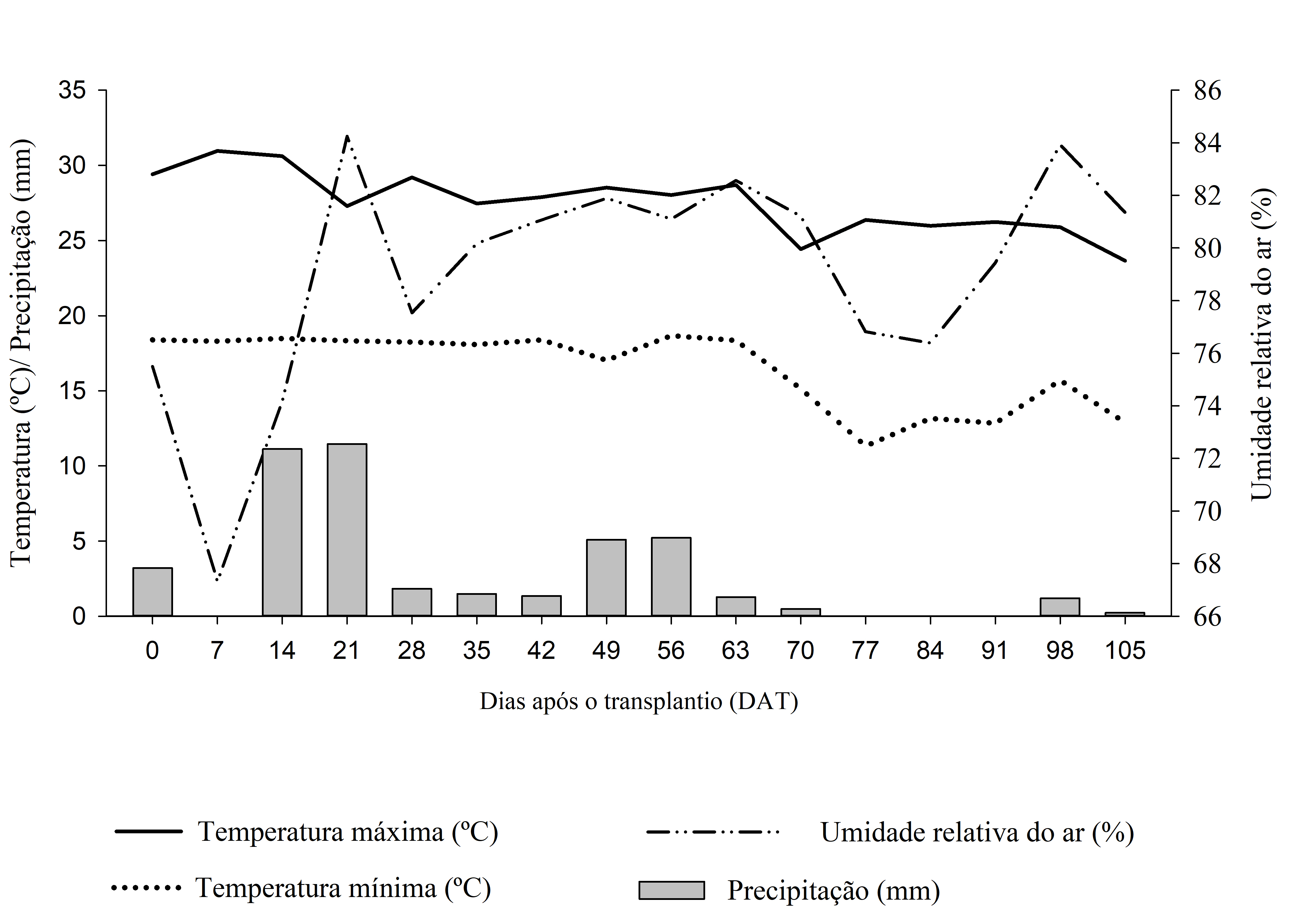
Figura 1: Médias semanais de temperatura máxima (ºC), temperatura mínima (ºC),
precipitação (milímetros) e umidade relativa do ar (%) observadas durante
a condução do experimento. (Dados cedidos pelo INMET).
Para enchimento dos vasos utilizou-se um solo de textura muito argilosa com as seguintes características químicas e físicas (Tabela 1).
Tabela 1. Características químicas e físicas do solo usado para enchimento dos vasos.
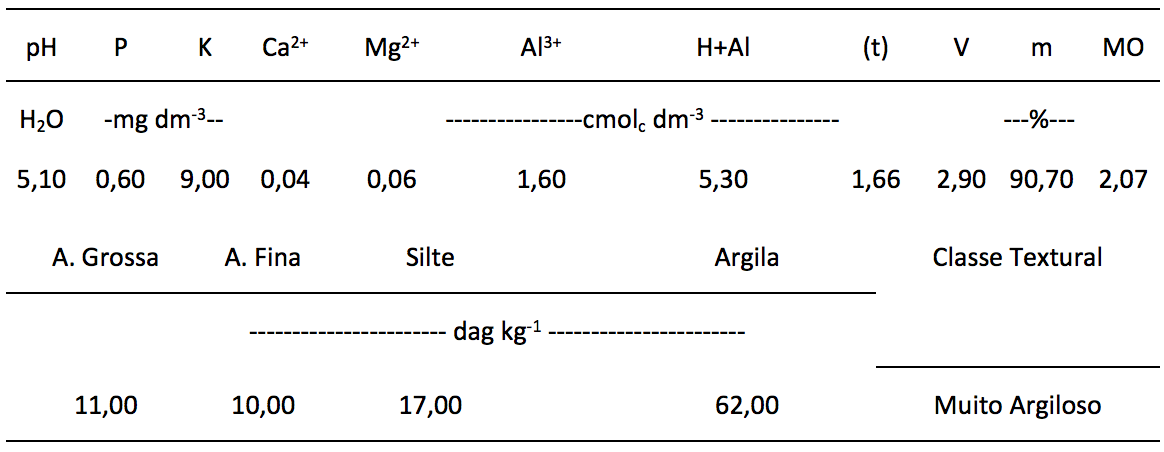
Análises realizadas no Laboratório de Análises de Solo Viçosa, segundo a metodologia da Empresa Brasileira de Pesquisa Agropecuária – EMBRAPA (1997); (t) = capacidade de troca catiônica efetiva; V = saturação por bases; m = Saturação por Al+3; MO = matéria orgânica.
Para adequação do substrato quanto à nutrição foram aplicados o equivalente a 1,5 t ha-1 de calcário dolomítico e 10,0; 0,45 e 0,38 kg m-3 de superfosfato simples, sulfato de amônio e cloreto de potássio, respectivamente.
Mudas padronizadas do clone AEC144, híbrido Eucalyptus urophylla x Eucalyptus grandis, com média de 23 cm de altura e três meses de idade, foram transplantadas no centro do vaso, simultaneamente, ao transplantio de três plantas de C. benghalensis com 4 a 6 folhas.
A adubação com micronutrientes nas doses de 10,0; 17,5 e 100,0 mg vaso-1, respectivamente de boro, cobre e zinco, foi realizada por via líquida e parcelada de duas vezes, aos 10 e 30 dias após o transplantio (DAT) das mudas de eucalipto.
As avaliações fisiológicas foram realizadas em duas épocas sendo a primeira avaliação realizada aos 30 dias e a segunda avaliação realizada aos 105 dias após o transplantio das mudas de trapoeraba.
As avaliações foram realizadas, no terço médio das plantas em uma folha nova de eucalipto, o que corresponde à quarta ou quinta folha completamente expandida contando do meristema apical para a base. Foi utilizado um analisador de gases no infravermelho (IRGA), marca ADC, modelo LCA 4 (Analytical Development Co. Ltd, Hoddesdon, UK), em local onde foi permitido livre circulação do ar. Cada bloco foi avaliado em um dia, entre 8 e 10 horas da manhã, de forma que mantivesse as condições ambientais homogêneas durante a avaliação.
As variáveis avaliadas foram consumo de CO2 (∆C - µmol mol-1), carbono interno (Ci - µmol mol-1), condutância estomática de vapores de água (Gs – mol m-1s-1), pressão de vapor na câmara subestomática e taxa de transpiração (E – mol H2O m-2s-1), taxa fotossintética (A - µmol m-2 s-1), sendo calculada ainda a eficiência do uso da água (UE – mol CO2 mol H2O-1) pela relação entre quantidade de CO2 fixado pela fotossíntese e quantidade de água transpirada.
Os dados obtidos foram transformados em porcentagem em relação à testemunha. Os resultados foram apresentados na forma de gráficos com desvio padrão.
Plantas de eucalipto em convivência com C. benghalensis mostram decréscimo nos valores da porcentagem da taxa fotossintética (A%) nas duas avaliações realizadas, sendo que, na segunda avaliação, com as plantas mais velhas e com maiores tempos de convivência esse decréscimo nos valores médios de A% foi maior. Destacando-se que na primeira avaliação a redução média da A% foi de cerca de 20% da testemunha. Já na segunda avaliação, esse decréscimo foi de 60% no maior período de convivência da planta daninha com a espécie arbórea (Figura 2).
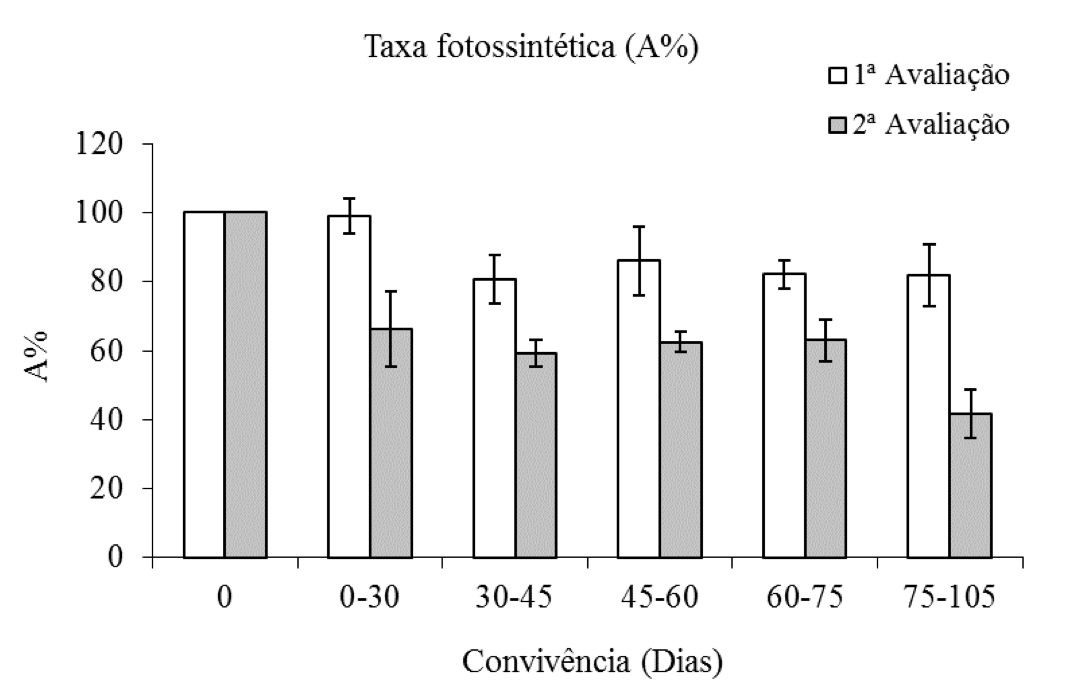
Figura 2: Porcentagem da taxa fotossintética de plantas de eucalipto (Clone AEC 144) – A%, em diferentes períodos de convivência com C. benghalensis. Médias representadas por barras e desvio-padrão. Taxa fotossintética da testemunha (1ª avaliação = 25,23 µmol m-2s-1; 2ª avaliação = 17,00 µmol m-2s-1).
Na primeira avaliação, em média a Gs% foi aproximadamente 40% menor em relação à testemunha, cultivada na ausência de competição (Figura 3). Entretanto, na segunda avaliação a porcentagem de condutância estomática (Gs%) do eucalipto apresentou incremento com o aumento do tempo de convivência com a planta daninha, sendo que, no maior período de convivência o valor de Gs% foi o dobro do observado na testemunha.
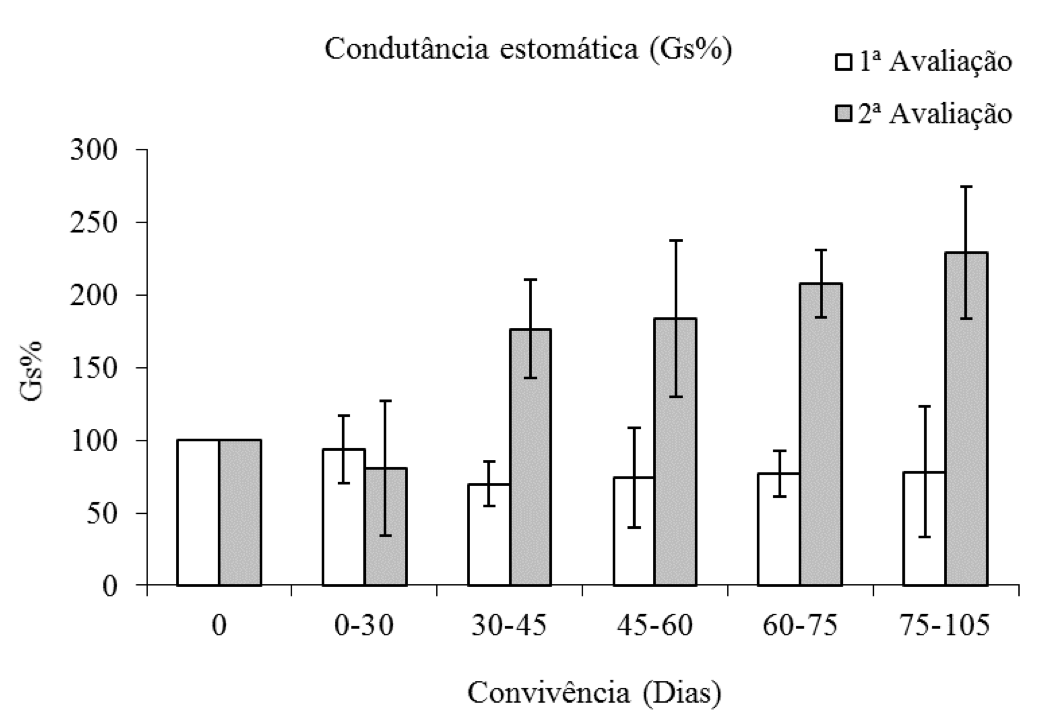
Figura 3: Porcentagem da condutância estomática de plantas de eucalipto (Clone AEC 144) – Gs%,
em diferentes períodos de convivência com C. benghalensis. Médias representadas por barras e desvio-padrão.
Condutância estomática da testemunha (1ª avaliação = 0,55 mol m-1 s-1; 2ª avaliação = 0,29 mol m-1 s-1).
Com relação à porcentagem de carbono interno (Ci%) na primeira avaliação, os tratamentos não diferiram da testemunha sem convivência com C. benghalensis. Na segunda avaliação, verificou-se acréscimo nos valores médios da Ci%, a partir dos 45 dias de convivência com C. benghalensis, no entanto, não foi constatada diferença entre os períodos de convivência com a espécie daninha (Figura 4).
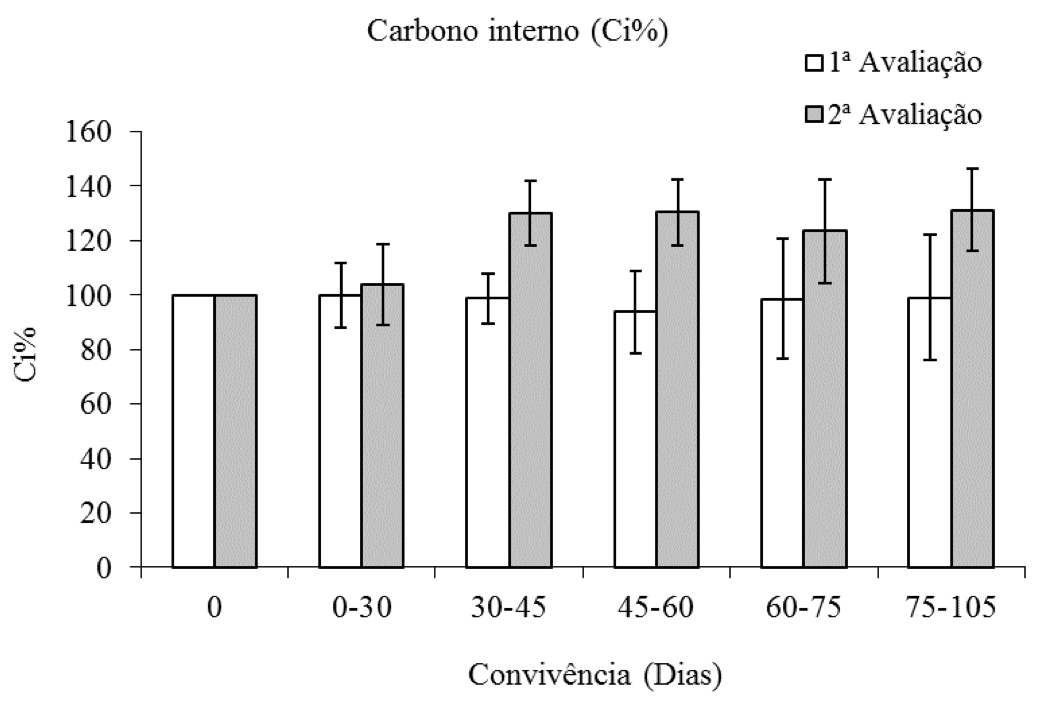
Figura 4: Porcentagem de carbono interno de plantas de eucalipto (Clone AEC 144) – Ci%,
em diferentes períodos de convivência com C. benghalensis Médias representadas por barras e desvio-padrão.
Carbono interno da testemunha (1ª avaliação = 262,71 µmol mol-1; 2ª avaliação = 305,23 µmol mol-1).
Na primeira avaliação a porcentagem da taxa transpiratória (E%) das plantas de eucalipto foi reduzida pela competição com a C. benghalensis. Já na segunda avaliação constatou-se aumento nos valores médios da E% do eucalipto dos 45 a 105 dias de convivência entre a espécie arbórea e a planta daninha (Figura 5).
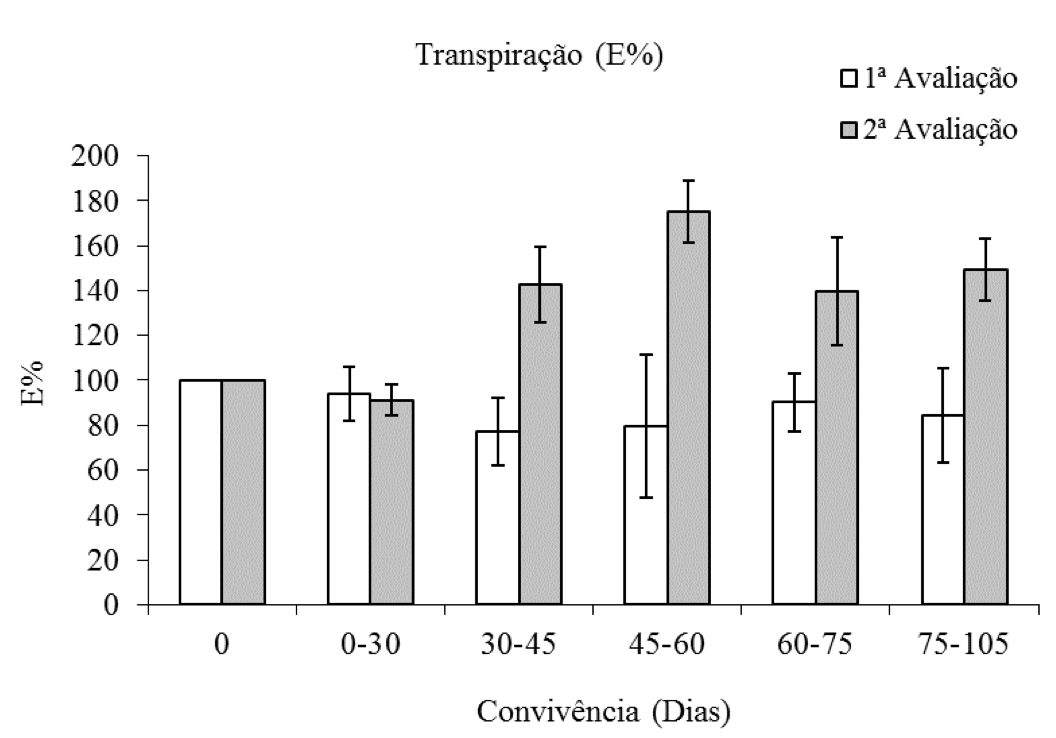
Figura 5: Porcentagem da taxa transpiratória de plantas de eucalipto (Clone AEC 144) – E%,
em diferentes períodos de convivência com C. benghalensis. Médias representadas por barras e desvio-padrão.
Taxa transpiratória da testemunha (1ª avaliação = 7,66 mol H2O m-2 s-1; 2ª avaliação = 3,00 mol H2O m-2 s-1).
Na primeira avaliação a porcentagem do uso da água (EA%) da espécie arbórea sem competição, não variou em relação às plantas em competição, no entanto, na segunda avaliação a redução da EA% das plantas de eucalipto foi de aproximadamente 70% dos 45 aos 105 dias de convivência do eucalipto com C. benghalensis (Figura 6).
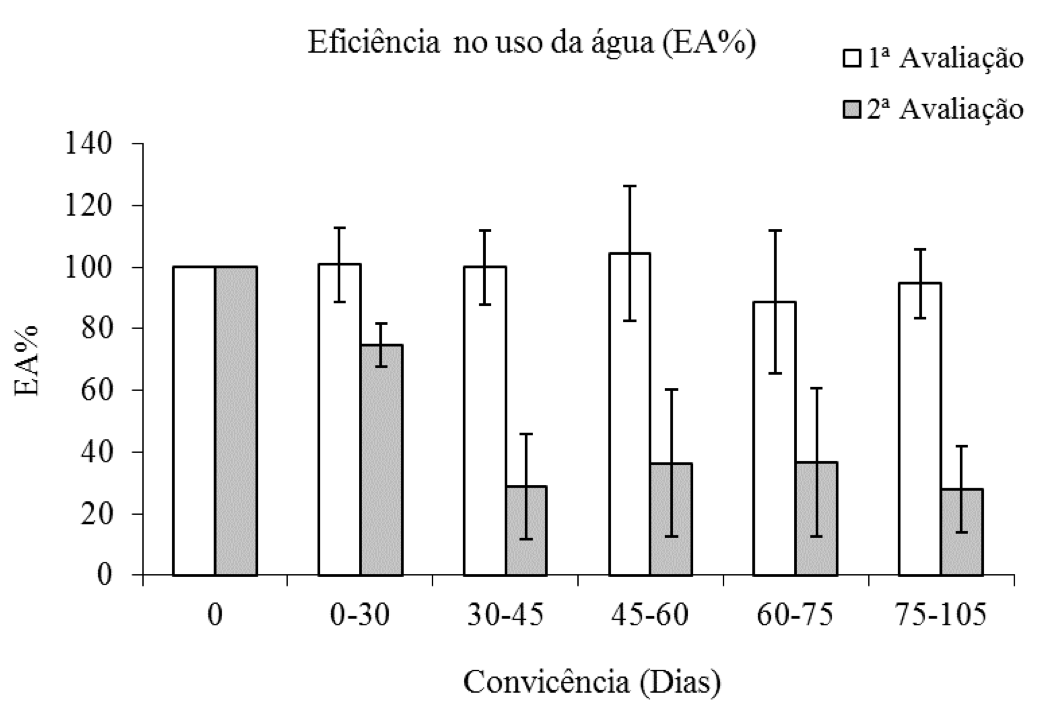
Figura 6: Porcentagem da eficiência no uso da água de plantas de eucalipto (Clone AEC 144) – EA%,
em diferentes períodos de convivência com C. benghalensis. Médias representadas por barras e desvio-padrão.
Eficiência no uso da água da testemunha (1ª avaliação = 3,46 μmol CO2 mmol-1 H2O;
2ª avaliação = 5,76 μmol CO2 mmol-1 H2O).
Com relação à porcentagem de diferença de temperatura (DT%) medidas em plantas de eucalipto, verificou-se que na primeira avaliação não foi observada diferença entre o eucalipto com e sem a convivência a com C. benghalensis, exceto aos 60 dias, período no qual os valores da DT% foram inferiores aos observados nos demais tratamentos. Na segunda avaliação constatou-se queda acentuada nos valores médios da DT%, reduções essas, superiores a cerca de 90% a partir dos 60 dias de convivência da planta daninha com a espécie arbórea (Figura 7).
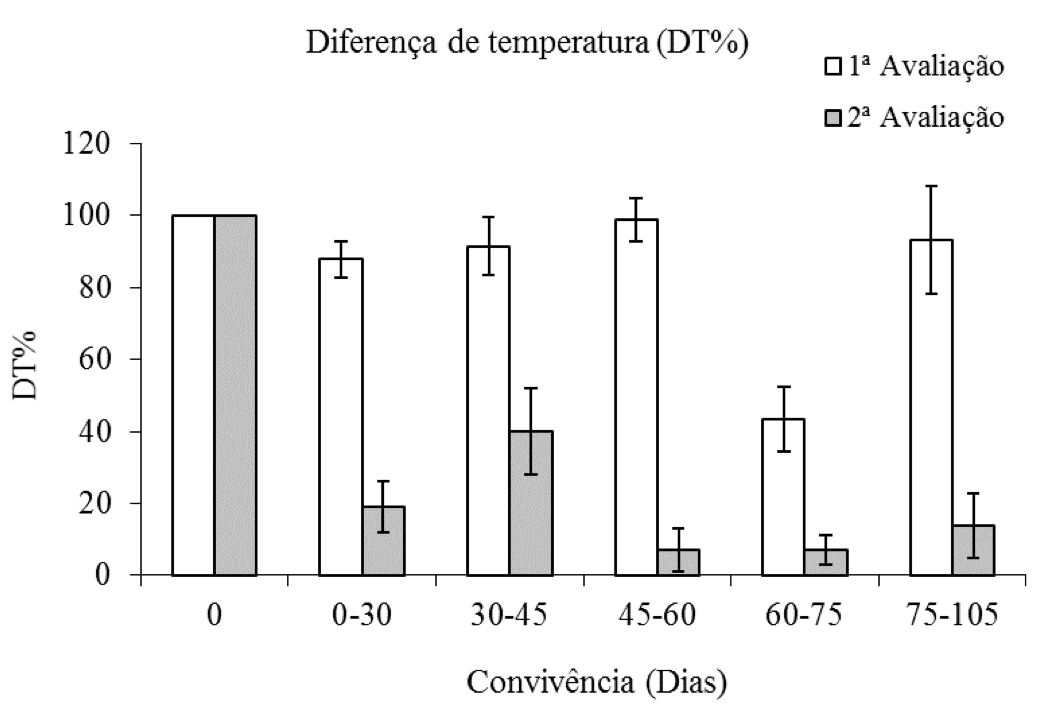
Figura 7: Porcentagem da diferença de temperatura de plantas de eucalipto (Clone AEC 144) – DT%,
em diferentes períodos de convivência com C. benghalensis. Médias representadas por barras e desvio-padrão.
Diferença de temperatura carbono atmosférica – carbono interno da testemunha
(1ª avaliação = 1,20 oC; 2ª avaliação = 1,60 oC).
A porcentagem da taxa de crescimento (TC%) do eucalipto mostrou redução com o incremento do período de convivência do eucalipto com C. benghalensis, sendo que, a partir dos 60 dias, essa redução foi superior a 80% (Figura 8).
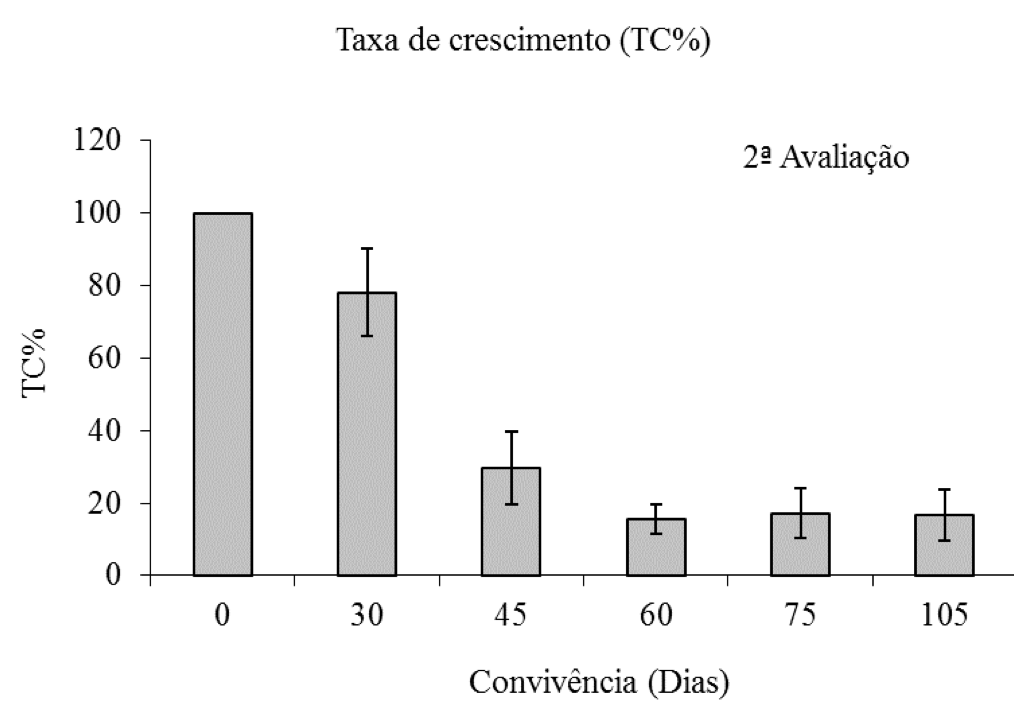
Figura 8: Porcentagem da taxa de crescimento de plantas de eucalipto (Clone AEC 144) – TC%,
em diferentes períodos de convivência com C. benghalensis. Médias representadas por barras e desvio-padrão.
Taxa de crescimento do eucalipto da testemunha (2ª avaliação = 3,02 g dia-1).
Na primeira avaliação da taxa fotossintética (A%) as plantas de eucalipto se encontravam em estádio mais jovem de desenvolvimento, sendo que, nessa fase as taxas fotossintéticas são maiores quando comparadas com plantas arbóreas em fases mais adiantadas de desenvolvimento (Taiz e Zeiger, 2013).
Manabe et al., (2014) trabalhando com a cultura do feijoeiro em competição com plantas daninhas, verificaram que o feijoeiro cultivado em competição com picão-preto apresentou menor taxa fotossintética do que o feijoeiro cultivado isoladamente. Matos et al., (2013) trabalhando com o efeito da competição de plantas de cobertura na entre linha da cultura do café, verificaram que a braquiária apresenta alta habilidade competitiva quando em maior densidade, reduzindo a taxa fotossintética, a condutância estomática e a taxa transpiratória do cafeeiro.
Reduções na taxa fotossintética quando uma planta está em estado de competição pode ser causada por déficit hídrico, deficiência de nutrientes e sombreamento proporcionado pelas plantas competidoras. As espécies arbóreas que crescem convivendo com as plantas daninhas podem apresentar deficiências de alguns nutrientes (Marchi et al., 1987), visto que a competição por nutrientes em povoamentos florestais pode impedir, especialmente em áreas recém implantadas, que as espécies expressem seu potencial de crescimento e produção.
Segundo Ronchi e Silva (2006), a redução dos teores nutricionais pode acarretar em reduções no crescimento e, por consequência, menor acúmulo de massa seca nas plantas, comprometendo a produtividade da cultura de café. A competição por nutrientes é um dos principais fatores ecológicos que alteram negativamente a produtividade das culturas agrícolas, assim como a competição por luz (Faustino, 2015).
A resposta mais significativa das plantas ao déficit hídrico consiste no fechamento dos estômatos. Quando as plantas são expostas a situações de déficit hídrico, exibem frequentemente respostas fisiológicas, que resultam de modo direto, na economia de água. Assim, a condutância estomática tem relação direta com a disponibilidade hídrica no solo (Bianchi et al., 2007).
A condutância foliar é composta em pequena parte pela condutância cuticular da epiderme e, quando os estômatos estão abertos, pela condutância estomática, que é controlada pelas células guarda dos estômatos. Assim, a condutância estomática é proporcional ao número e tamanho dos estômatos e aos diâmetros da abertura dos mesmos, características que dependem de outros fatores endógenos e ambientais (Brodribb e Holbrook, 2003).
A Ci é considerada variável fisiológica influenciada por fatores ambientais, como disponibilidade hídrica, de luz e energia, entre outros. O incremento na Ci pode indicar uma tentativa da planta de escapar do estresse gerado pela competição pelos recursos do ambiente (Matos et al., 2013). Dessa forma, na segunda avaliação as plantas de eucalipto encontravam-se em estádio mais avançado e sob período maior de competição com a planta daninha, o que pode explicar o maior acúmulo de carbono no interior da câmara subestomática.
A eficiência no uso da água é caracterizada como a quantidade de água transpirada por uma cultura para a produção de certa quantidade de matéria seca (Silva & Silva, 2013). Assim, culturas mais eficientes no uso da água podem produzir quantidade maior de matéria seca por grama de água transpirada. O uso mais eficiente da água está diretamente relacionado ao tempo de abertura estomática, pois, enquanto a planta absorve CO2 para a fotossíntese, a água é perdida por transpiração, com intensidade variável, dependendo de o gradiente de potencial entre a superfície foliar e a atmosfera, seguindo uma corrente de potenciais hídricos (Concenço et al., 2007).
O estudo da interferência da densidade de plantas daninhas em mudas de cafeeiro em outros trabalhos (Ronchi e Silva, 2006) evidenciaram redução severa da altura, número de folhas e biomassa seca da parte aérea das mudas de café em coexistência com C. diffusa, principalmente nas maiores densidades. Percebe-se que o aumento da densidade de algumas espécies daninhas intensifica a competição pelos recursos, afetando de forma mais severa o desenvolvimento da cultura. No presente estudo, o aumento do período de convivência da C. benghalensis com a espécie arbórea afetou de forma severa a taxa de crescimento do eucalipto.
A alteração na densidade de plantas, em uma determinada área, gera comportamento produtivo diferenciado, em função de competição por espaços, água, luz e nutrientes que se estabelece na comunidade vegetal (Zanine e Santos, 2004). Dessa forma, quando se misturam duas espécies são evidenciadas habilidades competitivas existentes entre ambas, sendo que a intensidade da interferência depende da habilidade dessas em competir pelos fatores de crescimento disponíveis no ambiente, e pela capacidade dessas espécies em produzir substâncias alelopáticas, além de outros fatores, fazendo predominar e/ou suprimir outra (Duarte et al., 2002).
Interessante destacar que o comportamento da A% e da TC% do eucalipto foi similar, sendo observado decréscimo nos valores das duas variáveis com o aumento do período de convivência da C. benghalensis com o eucalipto (Figuras 2 e 8). Segundo Concenço et al. (2008), a taxa fotossintética está diretamente relacionada com o consumo de CO2 do meio e com o aumento de massa das plantas.
Tarouco et al. (2009) trabalhando com eucalipto em competição com diversas espécies de plantas daninhas, verificaram que plantas de eucalipto que cresceram na ausência de competição apresentaram matéria seca total 400% superior a aquelas que conviveram com plantas de Lolium multiflorum, Brachiaria fasciculata, Cynodon dactylon, Digitaria horizontalis, Pennisetum clandestinum, Amaranthus lividus, Eupatorium buniifolium e Richardia brasiliensis. Faustino (2015) constatou que a convivência com C. benghalensis reduz o crescimento do eucalipto em altura, diâmetro do coleto, o acúmulo de matéria seca total, volume de raiz, área foliar e número de folhas.
A competição das plantas daninhas com as culturas leva ao menor fornecimento de alguns recursos para as espécies, ocasionando deficiências que culminam em alterações nas características fisiológicas relacionadas com a fotossíntese, como a deficiência hídrica (Floss, 2008) e nutricional (Melo et al., 2006) e a baixa qualidade ou quantidade de luminosidade (Concenço et al., 2009). Essas limitações podem levar a alterações na condutância estomática, na concentração interna de gases e, consequentemente, na atividade fotossintética e no uso eficiente da água.
O uso mais eficiente da água está diretamente relacionado ao tempo de abertura estomática, pois, enquanto o CO2 penetra na folha, a água é perdida pela transpiração, com intensidade variável, dependendo de o gradiente de potencial entre a superfície foliar e a atmosfera (Concenço et al., 2009).
O período de convivência com a C. benghalensis afeta as características fisiológicas das plantas de eucalipto, sendo mais intensa quando as plantas se encontram em estádios mais avançados de desenvolvimento e sobre maiores tempos de convivência com a C. benghalensis. Maior período de convivência da espécie arbórea com essa planta daninha prejudicam o crescimento da cultura. Os resultados dessa pesquisa reforçam a necessidade do controle da C. benghalensis em cultivos de eucalipto e indicam a necessidade de mais estudos que procurem compreender a interação entre culturas e plantas daninhas.
Bianchi, C. A. M., Bergonci, J. I., Bergamaschi, H., Dalmago, G. A., Heckler, B. M. M., Comiran, F. (2007) Condutância da folha em milho cultivado em plantio direto e convencional em diferentes disponibilidades hídricas. Ciência Rural, 37(2), 315-322.
Brodribb, T. J., e Holbrook, N. M. (2003) Stomatal closure during leaf dehydration, correlation with other leaf physiological traits. Plant Physiology, 132(4), 2166-2173.
Concenço, G., Ferreira, E. A., Silva, A. A., Ferreira, F. A., Galon, L., Reis, M. R., d'Antonino, L., Vargas, L., e Silva, L. V. B. D. (2008) Eficiência fotossintética de biótipos de azevém em condição de competição. Planta Daninha, 19(2), 247-253.
Concenço, G., Ferreira, E. A., Aspiazu, I., Silva, A. F., Galon, L., Ferreira, F. A., Silva, A. A., e Andres, A. (2009). Uso da água por plantas de arroz em competição com biótipos de Echinochloa crusgalli resistente e suscetível ao herbicida quinclorac. Planta Daninha, 27(2), 249-256.
Concenço, G., Ferreira, E. A., Silva, A. A., Ferreira, F. A., Viana, R. G., d’Antonino, L., Vargas, L. e Fialho, C. M. T. (2007) Uso da água em biótipos de azevém (Lolium multiflorum) em condição de competição. Planta Daninha, 25(3), 449-455.
Costa, N. V., Cardoso, L. A., Rodrigues, A. C. P., e Martins, D. (2008) Períodos de interferência de uma comunidade de plantas daninhas na cultura da batata. Planta Daninha, 26(1), 83-91.
Cruz, M. B., Alves, P. L. C. A., Karam, D., e Ferraudo, A. S. (2010) Capim-colonião e seus efeitos sobre o crescimento inicial de clones de Eucalyptus × urograndis. Ciência Florestal, 20(3), 391- 401.
Duarte, N. F., Silva, J. B., e Souza, I. F. (2002) Competição de plantas daninhas com a cultura do milho no município de Ijaci, MG. Ciência e Agrotecnologia, 26(5), 983-992.
Faustino, L. A. (2015) Convivência de eucalipto com Commelina benghalensis. (Dissertação de Mestrado). Universidade Federal de Viçosa. Viçosa.
Fleck, N. G., Bianchi, M. A., Rizzardi, M. A., e Agostinetto, D. (2006) Interferência de Raphanus sativus sobre cultivares de soja durante a fase vegetativa de desenvolvimento da cultura. Planta Daninha, 24(3), 425-434.
Floss, E. L. (2008) Fisiologia das plantas cultivadas. Passo Fundo: UPF.
Galon, L., Agostinetto, D., Moraes, P. V. D., Dal Magro, T., Panozzo, L. E., Brandolt, R. R. e Santos, L. S. (2007) Níveis de dano econômico para decisão de controle de capim-arroz (Echinochloa spp.) em arroz irrigado (Oryza sativa). Planta Daninha, 25(4), 709-718.
IBA - Indústria Brasileira de Árvores. Indicadores de desempenho do setor nacional de árvores plantadas: ano base 2014. 2016. Disponível em: <www.iba.org/shared/iba_2015.pdf>. Acesso em: 29 março. 2016.
Manabe, P. M. S., Matos, C. C., Ferreira, E. A., Silva, A. A., Sediyama, T., Manabe, A., Silva, A. F., Rocha, P. R. R., e Galon, L. (2014) Características fisiológicas de feijoeiro em competição com plantas daninhas. Biosciense Journal, 30(6), 1721-1728.
Marchi, S. R. (1987) Efeitos de períodos de controle das plantas daninhas no crescimento inicial e composição mineral de Eucalyptus grandis Hill ex Maiden. (Dissertação de Mestrado). Universidade Estadual Paulista "Júlio de Mesquita Filho". Jaboticabal.
Matos, C. C. et al. FIALHO, C. M. T., Evander Alves FERREIRA, E. A., SILVA, D. V., SILVA, A. A., SANTOS, J. B., França, A. C., GALON, L. (2013) Características fisiológicas do cafeeiro em competição com plantas daninhas. Biosciense Journal, 29(5), 1111-1119.
Melo, P. T. B. S., Schuch, l. O. B., Assis, F. N., e Concenço, G. (2006) Comportamento de populações de arroz irrigado em função das proporções de plantas originadas de sementes de alta e baixa qualidade fisiológica. Revista Brasileira de Agrociência, 12(1), 37-43.
Procópio, S. O., Santos, J. B., Silva, A. A., Martinez, C. A., e Werlang, R. C. (2004) Características fisiológicas das culturas de soja e feijão e de três espécies de plantas daninhas. Planta Daninha, 22(2), 211-216.
Rigoli, R. P., Agostinetto, D., Schaedler, C. E., Dal Magro, T., Tironi, S. (2008) Habilidade competitiva relativa do trigo (Triticum aestivum) e m convivência com azevém (Lolium multiflorum) ou nabo (Raphanus raphanistrum). Planta Daninha, 26(1), 93-100.
Rocha, D. C., Rodella, R. A., e Martins, D. (2007) Caracterização morfológica de espécies de trapoeraba (Commelina spp.) utilizando a análise multivariada. Planta Daninha, 25(4), 671-678.
Rodrigues, A. C. P., Filho, S. I. B. S., Martins, D., Costa, N. V., Rocha, D. C., e Souza, G. S. F. (2010) Avaliação qualitativa e quantitativa na deposição de calda de pulverização em Commelina benghalensis. Planta Daninha, 28(2), 421-428.
Ronchi, C. P., e Silva, A. A. Effects of weed species competition on the growth of young coffee plants. Planta Daninha, 24(3), 415-423.
Silva, A. A., e Silva, J. F. (2013) Tópicos em manejo de plantas daninhas. Viçosa: Universidade Federal de Viçosa.
Silva, D. V., Santos, J. B., Cury, J. P., Carvalho, F. P., Silva, E. B., Fernandes, J. S. C., Ferreira, E. A. e Concenço, G. (2013) Competitive capacity of cassava with weeds: Implications on accumulation of dry matter. African Journal of Agricultural Research, 8(6), 525-531.
Silva, W. (1993) Tolerância de Eucalyptus spp. a herbicidas e a eficiência desses produtos no controle de plantas daninhas. (Dissertação de Mestrado). Universidade Federal de Viçosa. Viçosa.
Taiz, L., e Zeiger, E. (2013) Fisiologia vegetal. Porto Alegre: Artmed.
Tarouco, C. P., Agostinetto, D., Panozzo, L. E., Santos, L. S., Vignolo, G. K., e Ramos, L. O. O. (2009). Períodos de interferência de plantas daninhas na fase inicial de crescimento do eucalipto. Pesquisa Agropecuária Brasileira, 44(9), 1131-1137.
Vidal, R. A., Spader, V., Fleck, N. G., Merotto Jr., A. (2004). Nível de dano econômico de Brachiaria plantaginea na cultura de milho irrigado. Planta Daninha, 22(1), 63-69.
Webster, T. M., e Grey, T. L. (2008). Growth and reproduction of Bengal Dayflower (Commelina benghalensis) in response to drought stress. Weed Science, 56(4), 561-566.
Zanine, A. M., e Santos, E. M. (2004). Competição entre espécies de plantas – uma revisão. Revista da FZVA. 11(1), 10-30.
1. Departamento de Fitotecnia. Universidade Federal de Viçosa. Engenheiro agrônoma. E email de contato: lais_araujo8@hotmail.com
2. Departamento de Fitotecnia. Universidade Federal de Viçosa. Engenheiro agrônoma. E email de contato: elisa.gomes@ufv.br
3. Departamento de Fitotecnia. Universidade Federal de Viçosa. Engenheiro agrônomo e Ph.D. E email de contato: lroberto@ufv.br
4. Departamento de Fitotecnia. Universidade Federal de Viçosa. Engenheiro agrônomo e Ph.D. E email de contato: gustavogamp@hotmail.com
5. Departamento de Fitotecnia. Universidade Federal de Viçosa. Engenheiro agrônomo e Ph.D. E email de contato: evanderalves@gmail.com
6. Departamento de Fitotecnia. Universidade Federal de Viçosa. Engenheiro agrônomo e M.Sc. E email de contato: agr.guilhermequeiroz@yahoo.com.br
7. Departamento de Zootecnia. Universidade Federal de Viçosa. Zootecnista. E email de contato: dteixeiras21@gmail.com